INTRODUCCIÓN
Los bosques brindan múltiples
servicios ecosistémicos y contienen una gran proporción de la biodiversidad
terrestre de la Tierra. Proporcionan hábitats para muchos grupos taxonómicos
que incluyen aves, vertebrados, invertebrados y microbios, que se ven afectados
de diferentes maneras por los bosques actuales y pasadas prácticas de manejo.
El tipo y la intensidad del manejo forestal son principales impulsores de la
diversidad estructural e impactan la biodiversidad en los ecosistemas
forestales. Conservación y restauración de los ecosistemas forestales es una de
las principales tareas críticas para la protección de ecosistemas globales.
(Madrigal-González et al., 2020; Oettel & Lapin, 2021; Toraño et al.,
2018). Al abordar la crisis mundial de la biodiversidad del siglo XXI, los
recientes esfuerzos políticos a nivel internacional, regional y local se han
centrado en los bosques, la biodiversidad y la contribución que puede hacer a
una biodiversidad más amplia metas de conservación. Por ejemplo, el Plan Estratégico
de las Naciones Unidas para los Bosques 2030 y su meta del Objetivo Forestal
Mundial 2, subraya la contribución de todos los tipos de bosques a la
conservación de la biodiversidad (UN, 2017). El logro de las metas mundiales de
diversidad biológica en los ecosistemas forestales gestionados requiere métodos
técnicamente apropiados, económicamente viables y socialmente aceptables con
criterios e indicadores bien definidos para ordenación forestal sostenible a
nivel local, regional y nacional (FAO, 2020).
La definición de diversidad de
especies considera tanto el número de especies; como el número de individuos o
abundancia de cada especie existente en un lugar determinado. Se evalúa
mediante índices, que son herramientas utilizadas en estudios florísticos y
ecológicos para comparar la diversidad de especies, ya sea entre tipos de
hábitat, tipos de bosques, etc., siendo los Andes el punto de biodiversidad más
rico de especies en el mundo. En este sentido, es necesario realizar
inventarios de vegetación para comprender la diversidad y el estado de
conservación en el que se encuentran a fin de complementar los esfuerzos para
reducir la incertidumbre sobre el conocimiento de la biodiversidad en estos
ecosistemas (Feng et al., 2021; Khaniya & Shrestha, 2020).
La diversidad de especies y su
estructura son esenciales para la biodiversidad forestal porque los árboles
proporcionan la necesidades básicas y hábitat de otras especies. Se ha aceptado
ampliamente que la distribución de especies y la estructura y su respuesta a
los factores ambientales son conceptos centrales para el estudio ecológico.
Además, la comprensión de la diversidad y la estructura de los bosques es
fundamental para el clima, que suele cambiar debido a la regulación, porque su
manipulación puede permitir la creación de bosques que absorban más carbono
dióxido. Por lo tanto, es importante que los bosques sean monitoreados
constantemente para determinar si se mantiene la diversidad. Sin embargo,
aunque se ha prestado mucha atención a la biodiversidad, su conservación y cómo
varía la biodiversidad entre las diferentes condiciones del bosque, todavía
solo tenemos una comprensión limitada de cómo la diversidad y la estructura de
los bosques varían entre las diferentes regiones de precipitación porque esos bosques
siempre se encuentran en diferentes regiones con diferentes climas (Cirimwami
et al., 2019; Eisawi et al., 2021; Khaine et al., 2017; Li et al., 2021)
La estructura del bosque también
puede mostrar una marcada variación entre continentes, como es el caso de los
bosques tropicales que son ricos en diversidad biológica y contienen una
proporción significativa de la diversidad biológica mundial, y varían
geográficamente dependiendo de la historia evolutiva y el clima. Por lo tanto,
el conocimiento de cómo la diversidad, distribución y estructura de los bosques
tropicales varía entre las regiones pluviales, siendo crítico para la ecología
tropical. Además, las relaciones entre especies y medio ambiente se pueden
utilizar como indicadores de las condiciones ambientales, la diversidad y los
patrones de los bosques se pueden utilizar para dilucidar la fitogeografía. Sin
embargo, a pesar de la importancia de esta información, se han obtenido pocos
datos detallados. obtenido para los bosques tropicales (Asuk et al., 2018;
Khaine et al., 2017). Se estima que la diversidad de especies vegetales y su
distribución en el espacio tienen efectos importantes sobre la función de los
ecosistemas de humedales. Sin embargo, el conocimiento de las relaciones entre
las especies de plantas y la diversidad espacial sigue siendo incompleto
(Brack, 2013; Chamané-Zapata et al., 2019).
Los Andes son el centro de
biodiversidad de especies más rico del mundo. La mayor parte de la
investigación y la atención de la conservación en los Andes se ha centrado en
biomas como el bosque lluvioso, el bosque nuboso y el páramo, donde la
diversidad de especies de plantas es el resultado de la rápida especiación
asociada con la orogenia andina reciente (Brack, 2013; Chamané-Zapata et al.,
2019).
El Perú es uno de los países más
valiosos de nuestro planeta, debido a su alta diversidad ecológica de climas,
franja altitudinal de vegetación y ecosistemas productivos. La alta diversidad
de ecosistemas ha permitido el desarrollo de numerosos grupos humanos. Se han
realizado estudios sobre las Poaceae en la región Huancavelica-Perú, en las
provincias de Huancavelica, Tayacaja y Castrovirreyna (Gutiérrez et al., 2019;
Gutiérrez & Castañeda, 2017); sin embargo, los ecosistemas forestales
altoandinos siguen siendo los menos estudiados y representan uno de los
sistemas más amenazados y menos gestionados (Chamané-Zapata et al., 2019).
Los bosques forestales, son
objeto de varios estudios para comprender mejor el papel que podrían desempeñar
en el desarrollo sostenible, el cambio climático y la biodiversidad florística.
Los bosques forestales, proporcionan muchos bienes y servicios ecosistémicos,
como la prevención de la erosión del suelo y la preservación de hábitats para
plantas y animales. El manejo sostenible de estos bosques requiere un buen
conocimiento de todos los árboles forestales naturales; este conocimiento sólo
podría ser confiable a través de estudios del medio forestal.
Los bosques están sometidos a una
gran presión antropogénica y requieren intervenciones de gestión para mantener
la biodiversidad, la productividad y la sostenibilidad en general. La
conservación de la biodiversidad a largo plazo depende básicamente del
conocimiento de la estructura, la diversidad de especies y las características
ecológicas de la vegetación. Los estudios de árboles forestales, son muy
importantes para el conocimiento de la diversidad de flora forestal, siendo
primordial la comprensión de los diversos aspectos ecológicos (Pauta, 2016;
Reátegui, 2015). Sin embargo, la explotación irracional de los árboles
forestales, se viene acentuando de gran manera como la tala y el tráfico
ilegal, motivo por el cual se ve mermado el uso potencial de la madera de los
bosques, especialmente de las especies de elevada valoración comercial.
Asimismo, las principales consecuencias de esta deforestación provoca la
pérdida de árboles empeorando el calentamiento global; el impacto en los
ecosistemas, debido a que los bosques preservan el agua, los suelos, las
plantas y la vida silvestre; pérdida de especies; daño al agua, pues los
bosques son represas naturales que atrapan el agua de lluvia en sus copas y en
las hojas, reteniendo y purificando el agua de lluvia y; la tala forestal
permite una rápida escorrentía y destruye la capacidad del suelo para absorber
agua (Keys et al., 2017; Pauta, 2016; Reátegui, 2015).
Por lo antes mencionado, la
presente investigación tiene como objetivo el estudio de la abundancia y
diversidad de árboles forestales de la comunidad andina de Ahuaycha, Tayacaja,
Huancavelica, siendo un instrumento vital para la obtención de información de
la flora forestal, permitiendo que los pobladores conozcan el potencial
forestal que cuentan, para su conservación, uso sostenible y sustentable de
dichos recursos.
METODOLOGÍA
Es un estudio descriptivo,
desarrollado en la Comunidad Andina de Ahuaycha, que se encuentra ubicada al
suroeste de la Provincia de Tayacaja en el Departamento de Huancavelica, a una
altura de 3262 m.s.n.m., abarcando una superficie forestal de 4.564 km2 y localizada
a 12°24'27'' de latitud sur y 74°'53'27'' de longitud oeste.
Las unidades de muestreo fueron
distribuidas de forma aleatoria, conformada por transectos de 20m x 50m, donde
se evaluaron todas las especies forestales con DAP (diámetro a la altura de
pecho) mayor a 10 cm. Registrándose la información recabada en campo de forma
separada por transecto (MINAM, 2015).
Para la determinación del cálculo
del tamaño del número de unidades de muestreo, se empleó la siguiente fórmula:
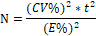
Donde:
N : Número de unidades de muestreo,
CV% : Coeficiente de variabilidad relacionado al volumen maderable
del bosque.
E% : Error de muestreo igual a 15 %.
T : 2, al 95 % de probabilidad.
Cabe indicar que se está
considerando el valor del CV% de 30%, para este tipo de bosques donde no se
cuenta con dicha información (MINAM, 2015).
Determinándose así el cálculo de
16 unidades de muestreo de 20m x 50m. (1000m2)
La composición de la flora de
árboles forestales fue representada por 7 especies identificadas en la
Comunidad Andina de Ahuaycha, Tayacaja, Huancavelica; distribuidas en cinco
familias: Cupressaceae, Myrtaceae y Pittosporaceae con 1 especies cada una; y, Pinaceae
y Rosaceae con 2 especies cada una (Tabla 2).
La especie más abundante fue Eucalyptus
globulus con 92.08%; y las especies con baja abundancia fueron Cupressus
macrocarpa, Pinus canariensis, Pinus radiata, Pittosporum tobira, Polylepis
incana y Prunus serótina, representando el 1.57%, 0.47%, 1.27%, 2.17%,
1.14% y 1.30% respectivamente (Tabla 3).
La especie con más frecuencia fue
Eucalyptus globulus con 30.19%; seguida de Pinus radiata
(18.87%), Pittosporum tobira (13.21%), Pinus canariensis (11.32%), Prunus
serótina (11.32%), Cupressus macrocarpa (9.43%) y Polylepis
incana (5.66%) (Tabla 3).
Los resultados de esta
investigación muestran que la Comunidad Andina de Ahuaycha presenta una riqueza
de 7 especies identificadas; se determinó el promedio de los índices de
diversidad de las 16 unidades de muestreo obteniéndose un índice de Simpson
(0.1561), Índice de Shannon – Wiener (0.3448) y el Índice de Margalef (0.4587),
representado todos estos índices una diversidad baja de especies de árboles
forestales (Tabla 4).
DISCUSIÓN
Los valores de abundancia y
diversidad de árboles forestales de la comunidad andina de Ahuaycha, aún
presenta un bajo nivel de alteración, debido a las diferentes actividades
antropogénicas desarrolladas en el área de influencia (Pauta, 2016; Reátegui, 2015).
La riqueza de las especies
locales y la variación de composición entre los valles altoandinos, difieren
entre los tipos de vegetación (Brandt et al., 2015; Flinn et al., 2008). Cabe
mencionar que la actividad del hombre ha cambiado la diversidad existente de la
flora forestal del mundo, siendo una señal de la recuperación de la flora
forestal después de haber sido utilizada (Walters et al., 2019).
Asimismo, se reportaron en Puno
para la provincia de Collao 45 especies de plantas en humedales altoandinos,
presentando una alta composición florística (Condori & Choquehuanca, 2001),
también en Bolivia, reportaron 58 especies de plantas en humedales altoandinos.
(Alzérreca et al., 2001). Sin embargo, en nuestro estudio se presentó una menor
composición de especies de árboles forestales en la comunidad andina de
Ahuaycha.
Exploraciones en el distrito de
Lircay, reportan un total de 46 especies y una subespecie de la familia
Poaceae, agrupadas en 21 géneros, 11 tribus y 6 subfamilias. El género Calamagrostis
es el más diverso con 9 especies, seguido de Poa con 5 especies. Asimismo, Aciachne
acicularis “paccupaccu”, Arundo donax “carrizo”, Cortaderia hieronymi y
Ortachne erectifolia “iruichu” son nuevos reportes para la región de
Huancavelica (Gutiérrez & Castañeda, 2014). En nuestro estudio en la
comunidad andina de Ahuaycha la familia Myrtaceae, es la más abundante con la
especie representante Eucalyptus globulus.
Estudios realizados en la
vegetación y composición florística de la microcuenca Pachachaca, ubicada en el
noroeste de Huancavelica, entre 2001 y 2003. Se registraron 180 especies
pertenecientes a 57 familias. La composición florística muestra una gran
riqueza en especies. Las Asteraceae fueron más representativas en las zonas
medias y bajas, Poaceae en las tierras altas y Fabaceae en las zonas medias y
bajas de la cuenca (Parra et al., 2004). Sin embargo, en nuestro estudio en la
comunidad andina de Ahuaycha se identificaron cinco familias: Cupressaceae,
Myrtaceae, Pittosporaceae, Pinaceae y Rosaceae, presentando menor composición
florística de árboles forestales.
También se logró reconocer 81
especies endémicas en 19 géneros. Las gramíneas endémicas del Perú se han
encontrado en prácticamente todas las regiones ecológicas reconocidas, aunque
la mayoría se encuentra en las regiones Puna Seca y Húmeda, Altoandina y
Mesoandina, desde el nivel del mar hasta los 5500 m de altitud. Se ha informado
de la existencia de veinticinco endemias en las áreas protegidas de Perú
(La-Torre et al., 2004). En comparación, en la comunidad andina de Ahuaycha
ubicada a una altitud de 3262m.s.n.m. se identificaron siete especies: Cupressus
macrocarpa, Eucalyptus globulus, Pinus canariensis, Pinus radiata, Pittosporum
tobira, Polylepis incana y Prunus serotine, presentando un bajo
número de especies.
Los resultados de los índices de
biodiversidad indican que la diversidad de vegetación de árboles forestales de
la comunidad andina de Ahuaycha presentan un bajo nivel de alteración, debido a
las diferentes actividades antropogénicas desarrolladas en el área de
influencia (ganadería, turismo) (Peralta-Peláez & Moreno-Casasola, 2009).
Además, en el estudio de la diversidad florística en pastizales según formación
vegetal en la subcuenca del río Shullcas, Junín, Perú informa que el índice de
Shannon-Wiener (H ') reveló que las formaciones vegetales en la parte alta
tienen una alta diversidad de 3,12 a 3,41; mientras que en la parte inferior
tienen una diversidad media de 2,75 y 2,81 (Yaranga et al., 2018). Estos
resultados son cercanos a los encontrados en los páramos, porque coincide con
el valor H 'más alto en el caso de pastizales con mayor cobertura (Caranqui et
al., 2016; Zheng et al., 2014). Este hecho reafirma la teoría de que los
ecosistemas de pastizales presentan diversidad heterogénea con la presencia de
diferentes especies entre ellos (Habel et al., 2013); esto indica que la
ubicación de las parcelas evaluadas en cada formación vegetal influye en el
índice obtenido (Habel et al., 2013). Sin embargo, los resultados de nuestro estudio
difieren, respecto a los bajos índices de diversidad de los árboles forestales
presentes de la comunidad andina de Ahuaycha.
Asimismo, cabe mencionar que en
la comparación de índices de diversidad se debe considerar las diferencias en
los métodos y áreas de muestreo. Por tanto, es necesario establecer los
elementos base y las condiciones bajo las cuales se deben calcular los índices,
de manera que aspectos como área de muestreo, hábito y categorías de tamaño,
que permitan realizar comparaciones entre ecosistemas similares (Chamané-Zapata
et al., 2019).
Los índices de diversidad son muy
útiles en la descripción de comunidades ecológicas. Dado que la diversidad en
una comunidad es expresión de la distribución de recursos y energía, su estudio
es uno de los enfoques más útiles en el análisis de comunidades (Rangel-Ch,
2015). Con respecto a la Riqueza de Especies (S) menciona que, la forma ideal
de medición, es tener un inventario completo que nos permita conocer el número
total de especies, obtenido por un censo de la comunidad (Rangel-Ch, 2015); sin
embargo, en la investigación este índice se ha determinado a partir de un
muestreo de las comunidades de vegetación de árboles forestales de las 16
unidades de muestreo en la superficie forestal de la comunidad andina de
Ahuaycha, basándose únicamente en el número de especies presentes en el área de
estudio.
CONCLUSIONES
La composición florística,
abundancia y frecuencia, indican que las familias más representativas de la
comunidad andina de Ahuaycha son: Cupressaceae, Myrtaceae, Pittosporaceae,
Pinaceae y Rosaceae.
La familia botánica con mayor
número de individuos fue Myrtaceae (2754 individuos). Las familias botánicas
con más especies fueron: Pinaceae (2 especies) y Rosaceae (2 especies).
La especie con mayor número de
individuos fue Eucalyptus globulus (2754), más abundante fue Eucalyptus
globulus con 92.08% y más frecuencia fue Eucalyptus globulus con
30.19%.
Según los índices de Simpson,
Índice de Shannon – Wiener y el Índice de Margalef, de los árboles forestales
la comunidad andina de Ahuaycha presenta una diversidad baja.
La información de la
biodiversidad florística obtenida, permite que los pobladores de la comunidad
andina de Ahuaycha conozcan el potencial forestal que cuentan, para su
conservación, uso sostenible y sustentable de dichos recursos.
REFERENCIAS BIBLIOGRÁFICAS
[1]
Alzérreca, H., Prieto, G., Laura, J., Luna, D., & Laguna, S. (2001).
Características y ditribución de los bofedales en el ambito boliviano
(Primera). ALT; PNUD. http://www.alt-perubolivia.org/Web_Bio/PROYECTO/Docum_bolivia/21.12.pdf
[2]
Asuk, S., Etim, E., Ifebueme, N., & Akpaso,
O. (2018). Species composition and diversity of mangrove swamp forest in
southern Nigeria. International International Journal of Avian & Wildlife
Biology, 3(2), 159–164. https://doi.org/10.15406/ijawb.2018.03.00078
[3]
Brack, A. (2013). Biodiversidad y Biocomercio en el Perú. In Meteorologische Zeitschrift (Vol. 22, Issue 2000).
[4]
Brako, L., & Zarucchi, J. (1993). Catalogue
of the flowering plants and gymnosperms of Peru. Missouri Botanical Garden.
EEUU.
[5]
Brandt, E., Petersen, J., Grossman, J., Allen,
G., & Benzing, D. (2015). Relationships between spatial metrics and plant
diversity in constructed freshwater wetlands. PLoS ONE, 10(8), 1–14.
https://doi.org/10.1371/journal.pone.0135917
[6]
Calderón, G., & Rzedowski, J. (2010). Flora fanerogámica del Valle
de México. In Instituto de Ecología, A.C (Edición di). Comisión Nacional para
el Conocimiento y Uso de la Biodiversidad. México, D.F.
[7]
Caranqui, J., Lozano, P., & Reyes, J. (2016). Composición y
diversidad florística de los páramos en la Reserva de Producción de Fauna
Chimborazo, Ecuador. Enfoque UTE, 7(1), 33–45.
https://doi.org/10.29019/enfoqueute.v7n1.86
[8]
Castellanos Bolaños, J., Treviño Garza, E., Aguirre Calderón, O.,
Jiménez Pérez, J., Musalem Santiago, M., & López Aguillón, R. (2008).
Estructura de bosques de pino pátula bajo manejo en Ixtlán de Juárez , Oaxaca.
Madera y Bosques, 14(2), 51–63. http://www.scielo.org.mx/scielo.php?script=sci_arttext&pid=S1405-04712008000200005
[9]
Chamané-Zapata, F., Custodio-Villanueva, M., Yaragua-Cano, R., &
Pantoja-Esquivel, R. (2019). Diversity of the riparian
vegetation of high Andean wetlands of the Junín region, Peru. Revista
Ambiente y Agua, 14(3), 1–15. https://doi.org/10.4136/1980-993X
[10] Cirimwami, L., Doumenge, C., Kahindo, J., & Amani, C. (2019). The
effect of elevation on species richness in tropical forests depends on the
considered lifeform: results from an East African mountain forest. Tropical
Ecology, 60(4), 473–484. https://doi.org/10.1007/s42965-019-00050-z
[11] Condori,
E., & Choquehuanca, D. (2001). Evaluación de las características y
distribución de los bofedales en el ámbito peruano del sistema TDPS (Primera). ALT;
PNUD.
https://docplayer.es/80123274-Evaluacion-de-las-caracteristicas-y-distribucion-de-los-bofedales-en-el-ambito-peruano-del-sistema-tdps.html
[12]
Eisawi, K., He, H., Shaheen, T., & Yasin, E.
(2021). Assessment of Tree Diversity and Abundance in Rashad Natural Reserved
Forest, South Kordofan, Sudan. Open Journal of Forestry, 11(01), 37–46.
https://doi.org/10.4236/ojf.2021.111003
[13] FAO. (2020). Sustainable forest management: lessons of history and
recent developments. Organización de las Naciones Unidas para la
Alimentación y la Agricultura. http://www.fao.org/forestry/sfm/es/
[14]
Feng, G., Huang, J., Xu, Y., Li, J., & Zang, R. (2021). Disentangling Environmental Effects on the Tree Species Abundance
Distribution and Richness in a Subtropical Forest. Frontiers in Plant Science,
12(1), 1–11. https://doi.org/10.3389/fpls.2021.622043
[15]
Flinn, K., Lechowicz, M., & Waterway, M.
(2008). Plant species diversity and composition of wetlands within an upland
forest. American Journal of Botany, 95(10), 1216–1224. https://doi.org/10.3732/ajb.0800098
[16] Font,
P. (2001). Diccionario de Botánica (Segunda Ed). Ediciones Península.
Barcelona. España.
[17] Gutiérrez,
H., & Castañeda, R. (2014). Diversidad De Las Gramíneas (Poaceae) De Lircay
(Angaraes, Huancavelica, Perú). Ecología Aplicada, 13(1), 23–33.
https://doi.org/10.21704/rea.v13i1-2.451
[18] Gutiérrez,
H., & Castañeda, R. (2017). Catálogo De Las Gramíneas (Poaceae) De
Huancavelica, Perú. Ecología Aplicada, 16(1), 63–73.
http://www.scielo.org.pe/pdf/ecol/v16n1/a08v16n1.pdf
[19]
Gutiérrez, H., Castañeda, R., & Nauray, W. (2019). Epidendrum suinii (Orchidaceae: Epidendroideae) a new record for
peruvian flora. Revista Peruana de Biologia, 26(2), 271–274.
https://doi.org/10.15381/rpb.v26i2.15142
[20]
Habel, J., Dengler, J., Janišová, M., Török, P.,
Wellstein, C., & Wiezik, M. (2013). European grassland ecosystems:
Threatened hotspots of biodiversity. Biodiversity and Conservation, 22(10),
2131–2138. https://doi.org/10.1007/s10531-013-0537-x
[21]
Keys, P., Wang-Erlandsson, L., Gordon, L.,
Galaz, V., & Ebbesson, J. (2017). Approaching moisture recycling
governance. Global Environmental Change, 45(1), 15–23.
https://doi.org/10.1016/j.gloenvcha.2017.04.007
[22]
Khaine, I., Woo, S., Kang, H., Kwak, M., Je, S.,
You, H., Lee, T., Jang, J., Lee, H., Lee, E., Yang, L., Kim, H., Lee, J., &
Kim, J. (2017). Species diversity, stand structure, and species distribution
across a precipitation gradient in tropical forests in Myanmar. Forests, 8(8),
1–15. https://doi.org/10.3390/f8080282
[23] Khaniya, L., & Shrestha, B. (2020). Forest regrowth reduces
richness and abundance of invasive alien plant species in community managed
Shorea robusta forests of central Nepal. Journal of Ecology and
Environment, 44(1), 1–8. https://doi.org/10.1186/s41610-020-00158-7
[24] La-Torre,
M., Cano, A., & Tovar, O. (2004). Las Poáceas del Parque Nacional
Yanachaga-Chemillén (Oxapampa, Perú). Parte II: Pooideae, Centothecoideae,
Arundinoideae, Chloridoideae y Panicoideae. Revista Peruana de Biologia, 11(1),
51–70. https://doi.org/10.15381/rpb.v11i1.2433
[25] Li,
S., Liu, W., Lang, X., Huang, X., & Su, J. (2021). Species
richness, not abundance, drives ecosystem multifunctionality in a subtropical
coniferous forest. Ecological Indicators, 120(1), 1–8.
https://doi.org/10.1016/j.ecolind.2020.106911
[26] López-Hernández,
J., Aguirre-Calderón, O., Alanís-Rodríguez, E., Monarrez-Gonzalez, J.,
González-Tagle, M., & Jiménez-Pérez, J. (2017). Composición and diversidad
de especies forestales en bosques templados de Puebla, México. Madera y
Bosques, 23(1), 39–51. https://doi.org/10.21829/myb.2017.2311518
[27]
Madrigal-González, J., Calatayud, J., Ballesteros-Cánovas, J.,
Escudero, A., Cayuela, L., Rueda, M., Ruiz-Benito, P., Herrero, A., Aponte, C.,
Sagardia, R., Plumptre, A., Dupire, S., Espinosa, C., Tutubalina, O., Myint,
M., Pataro, L., López-Sáez, J., Macía, M., Abegg, M., … Stoffel,
M. (2020). Climate reverses directionality in the richness–abundance
relationship across the World’s main forest biomes. Nature Communications,
11(1), 1–7. https://doi.org/10.1038/s41467-020-19460-y
[28]
Magurran, A. (2004). Measuring Biologcial
Diversity. In Blackwell Science Ltd (Ed.), Blackwell Publishing (First edit).
Blackwell Publishing.
http://www.bio-nica.info/Biblioteca/Magurran2004MeasuringBiological.pdf
[29] MINAM.
(2015). Guía de inventario de la flora y vegetación. In M. del Ambiente. (Ed.),
Ministerio del Ambiente (Primera ed). Dirección General de Evaluación,
Valoración y Financiamiento del Patrimonio Natural.
http://www.minam.gob.pe/patrimonio-natural/wp-content/uploads/sites/6/2013/10/GUÃ-A-DE-FLORA-Y-VEGETACIÓN.compressed.pdf
[30] Mostacero,
J., Castillo, F., Mejía, F., Gamarra, O., Charcape, J., & Ramírez, R.
(2011). Plantas Medicinales del Perú: Taxonomía, Ecogeografía, Fenología y
Etnobotánica (Proimera e). Asamblea Nacional de Rectores Fondo Editorial. Trujillo,
Perú.
[31]
Mostacero, J., Mejía, F., & Gamarra, O. (2009). Fanerógamas
del Perú. Taxonomía, utilidad y ecogeografía (Primera ed). Concytec.
Trujillo, Perú.
[32] Oettel, J., & Lapin, K. (2021). Linking forest management and
biodiversity indicators to strengthen sustainable forest management in Europe. Ecological
Indicators, 122(1), 1–13. https://doi.org/10.1016/j.ecolind.2020.107275
[33] Parra,
F., Torres, J., & Ceroni, A. (2004). COMPOSICIÓN FLORÍSTICA Y VEGETACIÓN DE
UNA MICROCUENCA ANDINA: EL PACHACHACA (HUANCAVELICA). Ecología Aplicada, 3(1),
9–16. http://www.scielo.org.pe/pdf/ecol/v3n1-2/a02v3n1-2.pdf
[34] Pauta,
L. (2016). Cálculo del índice de biodiversidad florística en el bosque
protector Aguarongo [Universidad Politécnica Salesiana].
https://dspace.ups.edu.ec/bitstream/123456789/11887/1/UPS-CT005645.pdf
[35] Peralta-Peláez,
L., & Moreno-Casasola, P. (2009). COMPOSICIÓN FLORÍSTICA Y DIVERSIDAD DE LA
VEGETACIÓN DE HUMEDALES EN LOS LAGOS INTERDUNARIOS DE VERACRUZ. Boletín de La
Sociedad Botánica de México, 85, 89–101.
https://www.redalyc.org/pdf/577/57712090009.pdf
[36] Rangel-Ch,
J. (2015). La biodiversidad de Colombia: significado y distribución regional.
Revista Académica de Colombia de Ciencias Experimentales, Físicas y Naturales,
39(151), 176–200. https://doi.org/10.1093/nar/16.9.3655
[37] Reátegui,
J. (2015). Estructura horizantal y diversidad florística de un bosque denso de
terrazas en áreas de perforanción del lote 174, Ucayali-Perú [Universidad
Nacional de la Amazonía Peruana]. In Universidad Nacional de la Amazonía
Peruana (Vol. 1).
http://repositorio.unapiquitos.edu.pe/bitstream/handle/UNAP/4477/Ida_Tesis_Titulo_2015.pdf?sequence=1&isAllowed=y%0Ahttps://fcf.unse.edu.ar/archivos/series-didacticas/SD-06.pdf
[38]
Shannon, C., & Weaver, W. (1964). The
Mathematical theory of communication (Champaign (ed.); Décima edi). University
of Illinois Press.
https://pure.mpg.de/rest/items/item_2383164_3/component/file_2383163/content
[39] Toraño, A., Biber, P., Poschenrieder, W., Schwaiger, F., &
Pretzsch, H. (2018). Forestry projections for species diversity-oriented
management: an example from Central Europe. Ecological Processes, 7(23), 1–18.
https://doi.org/10.1186/s13717-018-0135-7
[40] UN. (2017). Resolution adopted by the General Assembly on 27 April
2017. In United Nations (Ed.), United Nations strategic plan for forests (Vol.
35, Issue 2). United Nations strategic plan for forests 2017–2030.
[41] Vasquez,
R., & Rojas, R. (2016). Clave para identificar grupos de familias de
Gymnospermae y Angiospermae del Perú. Jardín Botánico de
Missouri. EEUU.
[42] Walters, G., Fraser, J., Picard, N., Hymas, O., & Fairhead, J.
(2019). Deciphering African tropical forest dynamics in the
Anthropocene : how social and historical sciences can elucidate forest
research and management. Anthropocene, 27(1), 1–12. https://serval.unil.ch/resource/serval:BIB_FF8380BAB22E.P001/REF
[43] Yaranga,
R., Custodio, M., Chanamé, F., & Pantoja, R. (2018). Diversidad florística
de pastizales según formación vegetal en la subcuenca del río Shullcas, Junín,
Perú. Scientia Agropecuaria, 9(4), 511–517.
https://doi.org/10.17268/sci.agropecu.2018.04.06
[44] Zheng, S., Li, W., Lan, Z., Ren, H., Wang, K., & Bai, Y. (2014).
Testing functional trait-based mechanisms underpinning plant responses to
grazing and linkages to ecosystem functioning in grasslands. Biogeosciences
Discussions, 11(9), 13157–13186. https://doi.org/10.5194/bgd-11-13157-2014